
Dynamics of Akt isoforms and role of Immune Evader (RCAS 1) in different grades of Breast Cancer tissues in Pakistani Women
Mahmood H. Qazi, Mohammad Javed Iqbal
Affiliation
University of Lahore, Lahore, Pakistan
Corresponding Author
Ahsan Sattar Sheikh, University of Lahore, Lahore- 54000, Pakistan; E-mail: assheikh@bzu.edu.pk
Citation
Sheikh, A.S., et al. Dynamics of Akt Isoforms and Role of Immune Evader (RCAS 1) in Different Grades of Breast Cancer Tissues in Pakistani Women. (2018) Int J Cancer Oncol 5(1): 16- 25.
Copy rights
© 2018 Sheikh, A.S. This is an Open access article distributed under the terms of Creative Commons Attribution 4.0 International License.
Keywords
Breast Cancer, Akt isoforms, RCAS 1, Invasion, Pakistan
Abstract
Breast Cancer (BC) is the leading cause of death among females. It has been documented that Akt isoforms and kRaS control the proliferative pathways in most cancer including BC. To address the issue of breast cancer signaling through mRNA quantification along with proteins quantifications in Akt pathway by microbead assay.
A total of 42 specimens were selected from a pool of samples representing nearly all Bloom’s Richardson’s grades tissues (Normal, Hyperplasia, Ductal in situ Carcinoma, Grade I, Grade II and Grade III). No Grade IV sample was available. Total of 8 genes (Akt 1, Akt 2, Akt 3, p53, RCAS 1, Bcl 2, BclxL, and Mcl 1) and 11 proteins in PI3K / Akt - mToR pathway were assayed simultaneously using microbead BioPlex™ assay. Results on quantification of RCAS 1 gene (Immune Evader) indicates its significance by increased quantities to help to evade the breast cancer cells to distant places to metastasize in the body. Data obtained on Akt isoforms (Akt 1, Akt 2, Akt 3) show that all three play significant role in the progression of breast cancer. Our data indicate that in advance grades (grade II and III) Akt 3 along with Akt 1 act as oncogenes. Here we propose that Akt1 and Akt 3 appear to be involved in cellular growth/proliferation and angiogenesis/invasion respectively. Bioinformatics modeling have shown that the conformation of Akt 3 differs from Akt 1 and Akt 2 in PH domain that interacts to surface membrane from inner side.
Based on our data, we conceived that roles of all three Akt isoforms in growth, hypertrophy, epithelial mesenchymal transition (EMT), adhesion, and metastasis been highlighted. As reported in other cancers, Akt 3 appears to be involved in the phosphorylation of VEGF and Integrin family proteins, thereby facilitating invasion and metastasis in the aggressive stages of breast cancer. We propose a model with dynamics of Akt and RCAS 1 for metastasis.
Introduction
In this part of world (Pakistan), breast cancer is the second most common cancer after lung cancer and is ranked first in women[1-4]. The current information indicates that breast cancer is most common in young women of Pakistan[5-9]. It has been substantiated that Akt and kRaS-MAPK are dominant proliferative pathways stimulated by activated Receptor Tyrosine kinase (RTK)[10,11]. Akt (also termed Protein Kinase B – PKB) works as double edged sword for cell survival as well as inhibitor of apoptosis[12,13]. It has been identified that Akt, especially Akt 1 isoform is the principle oncogene[7]. More recently, three isoforms namely, Akt 1, Akt 2, Akt 3 have been reported[1,2]. The function of each isoform has yet to be fully substantiated. So far three corresponding Akt genes (Akt 1, Akt 2 and Akt 3) have been reported[14-17]. The differences in functional abnormalities observed in Akt null mice suggest that all isoforms have physiologically diverse roles[18]. Characterization of Akt 1 (Ch 14q32.33), the prototype isoform, makes it the key signaling molecule in diverse cellular processes including muscle hypertrophy[19-21] and presented as a principle proliferative molecule in many types of cancer[22,23]. Akt 2 (Ch 19q13.2) in contrast to Akt 1, knockout results in mild growth deficiency with severe diabetic phenotype (insulin resistance)[23,24]. The function of Akt 3 (Ch 1q44), a comparatively less explored isoform, is predominantly associated with brain growth and testes[20,25,26]. The Akt isoforms are ~480 amino acids long protein with a large number of posttranslational phosphorylation sites[10,27]. Initial records indicate that Akt 3 is consistently detected in cancer tissues as compared to normal tissues. However, this remains controversial[28,29]. Furthermore, increase in quantities of RCAS 1 seems to be associated with release of Akt 3 from Golgi complex[30-32]. RCAS 1 is over expressed in many patients suffering from breast carcinoma and its expression levels correlate with tumor grades as well, suggesting that it may be involved in immune escape[31,33]. RCAS 1 is also been implicated in regulatory functions with VEGF in ovarian cell line[34,35].
Bcl 2 family genes / proteins maintain a critical balance between cell proliferation and apoptosis[36,37]. These antiapoptotic proteins include Bcl 2, BclxL and Mcl 1. These proteins have a common hydrophobic groove formed by BH 1, BH 2 and BH 3 domains[38-40]. Evidence is also available for the overexpression of some members of the Bcl 2 family in breast cancer[36,40]. The tumorigenic potential of these proteins has been well documented in animal models in which expression of Bcl 2 oncogene has been reported in a variety of tumors and in lymphomas[41,42]. High expression of Bcl 2 has been observed in Estrogen Receptor (ER) and Progesterone Receptor (PR) positive breast cancer[43,44]. BclxL on the other hand is related to the invasion and metastasis of some solid tumors[40,45,46]. Mcl 1 is shown to be regulated by mToR complex for transcription and translation[47,48]. Significance of Mcl 1 expression has been studied in a number of tumor types. It is linked to poor patient prognosis and survival[49].
The three isoforms have been studied in melanoma[14,50,51]. In melanoma there is sharp rise in Akt 3[52,53]. This aspect has, however, not been studied adequately in other cancers, including breast cancer. Addition to it, little information is available on the molecular pathogenesis of Akt 1, Akt 2 and Akt 3 in various grades of cancer. Literature doesn’t provide concurrent role of the three isoforms in breast cancer. Lack of this information about the dynamics of Akt isoforms has prompted us to examine the amplification of Akt 1, Akt 2 and Akt 3 in normal, GradeI, Grade II and Grade III tissues supported by assay of 11 phosphoproteins of Akt pathway. This will be first report on the dynamics of Akt isoforms and their associated substrates in Pakistani (Punjabi) population.
Materials and Methods
Specimen Collection and Processing
Total of 42 breast cancer specimens in different Bloom Richardson’s grades[28] were collected from three major public and private hospitals of Lahore, Pakistan. The specimens were stored at – 40°C (Forma Scientific, USA) that includes Formalin Fixed Paraffin Embedded (FFPE) tissue blocks and Formalin Fixed (FF) tissues. The tissues were grounded to powder form in thistle and mortar with liquid nitrogen (Fine Gas (Pvt) Ltd, Lahore). Materials were taken out when ever needed on crushed ice bucket (Zellegra Eismaschinen, Germany) for further processing.
RNA Isolation, cDNA, Primer Design and qPCR Quantification
Total RNA was extracted by LiCl – Urea method (3M LiCl, 6M Urea, 50 mM Tris (7.4), 1 mM EDTA Na2, 0.5% Sarkosyl (Serva, Germany) to preweighed tissue specimen (40-60 mg)[54]. Total of 15 – 20 μL of 10% SDS was added to LiCl – Urea reagent to enhance homogenization with Micropestle (Bel Air Products, USA) two times for 5 minutes each. Total RNA was extracted from cells treated with Proteinase K (Vivantis, Malaysia) in PK buffer (10 mM Tris (8.0), 2 mM EDTANa2 (8.0), 200 mM NaCl, 0.5% SDS and 200 μg / mL Proteinase K (20 mg/mL) for 30 minutes at 37°C. Samples were Phenol – Chloroform – Isoamyl alcohol (PCI) extracted (25:24:1) (MP Bio, USA) twice and with chloroform (Merck, Germany) once[55]. The aqueous phase was precipitated with HPLC grade 2-Propanol (BDH, UK) over night at – 40°C[56]. A second precipitation was done with ½ volume Ammonium acetate (5 M) and 2.5 volume of molecular grade ethanol (Merck, Germany) after pellet been dissolved in 100 μL of RNase free water. Sample was centrifuged (Sigma, Germany) at 17 kg for 20 minutes at 4°C, pallets washed with 70% RNase free ethanol, air dried and then at 45°C for 10 minutes on heating block (T-box, Jena Analytika, Germany).
Pellets were dissolved in molecular grade water and RNA concentration was adjusted to 1 μg/μL (Jena Analytika, Germany) used for cDNA synthesis by setting up a RT reaction with Maxime™ RT Random Hexamer kit (Inron Biotechnology, S. Korea) in 20 μL volumes. The RT PCR program was written on Thermocycler (BioRad C1000 with CFX96 detector, USA) with annealing temperature of primers at 16°C (10 minutes), 25°C (10 minutes), 37°C (40 minutes) and final elongation at 42°C for 20 minutes. Reaction was denatured at 95°C for 5 minutes. Multiplex real time (rt) quantification were done with gene specific primers for Akt 1 (AlexaFluor 647), Akt 2 (JOE), Akt 3 (Cy 5), p53 (FAM) with GAPDH (ROX) as housekeeping normalization. Primers were designed from FASTA sequences uploaded on to either Light Upon eXtension (LUX™) primer design software; D – LUX primer design software (Invitrogen Corp., USA) or QuantPrime (Max Planck Institute, Germany). Labeled primers with fluorescent tags were synthesized from BioBasicInc, USA or IDT, USA. Primer sequences are given Table 1 and Table 2. All qPCRs were done by using qMaster Mix with UDG (Invitrogen, USA). Total of 2 μL cDNA reaction, 400 nmol of each fluorescent labeled primer were used in each 20 μL reactions. Amplification protocol was initiated with heating at 52°C (2 minutes), denaturation at 95°C (10 minutes), cycle denaturation at 95°C (20 seconds), annealing at 58°C (30 seconds), and elongation at 72°C (30 seconds). The fluorescent signal was collected for 45 cycles. The CFX manager (BioRad, USA) was used to optimize the quantified data. Each quantification reaction was done three times in separate experiments.
Micro Bead Milliplex MAP 11-plex Assay
Akt / mToR Panel (Milliplex, USA) was selected for 19 selected samples with traditional Bioplex / Luminexx MAP based platform to detect simultaneously multiple protein quantities. Each sample was weighed ~50 mg in sterile, oven dried microfuge tube for protein isolation. Assay was run according to the manufacturer manual’s protocol provided with the Akt – mToR panel kit. Before the processing of each sample, tissue powder was washed in 0.5 mL ice cold TBS and to pellet 150 μL ice cold 1X Milliplex MAP Lysis Buffer containing freshly prepared 1X protease inhibitors and 1X phospho inhibitors cocktails (Serva, Germany). Gently rock the lysate for 10 - 15 minutes at 4°C. Remove particulate matter by high speed centrifugation at 15 kg. Aliquot clarified supernatant and store the lysate at –40°C. Lysate protein concentrations were determined by diluting 1:10 in PBS by Qubit™ (Invitrogen, USA) protein assay.
Statistical Data Analysis
All genes and protein quantifications were statistically analyzed by SPSS 17 (SPSS, Inc. USA) using various tools in the software.
Bioinformatics Tools: Different bioinformatics tools were used to get 3D protein structures of different molecules which includes; Akt 1, Akt 2, Akt 3 by loading protein FASTA Sequences on to LOMETS software (http://zhanglab.ccmb.med.umich.edu/LOMETS). The molecules were displayed by PyMol, Version 1.3 (2010).
Results
Physical features of isoforms
In the Figure 1a that represents the conserved domains of the three isoforms. The same figure also shows the percentage of homology between three Akt isoforms. Pleckstrin Homology (PH) domain has the highest homology percentage between the three isoforms[29]. The highest percentage of homology is observed between Akt 1 and Akt 3 (84%), followed by 80% between Akt 1 and Akt 2. There is only 76% homology between Akt 2 and Akt 3. Variations in other domains have also been recorded (Figure 1a).

The primary sequence was down loaded in FASTA format and protein alignment was done with MAFTT tool. Serine, threonine and tyrosine amino acids were highlighted with; S Serine T Threonine Y Tyrosine in the aligned amino acid sequence. The box represents a hyper variable region in the kinase-regulatory region where Y452 is identified as unique addition. These amino acids are labeled in the LOMETS 3D prediction software. All 3D data is visualized in PyMol. The highlighted box represents the variable region in regulatory linker region of the three Akt isoforms
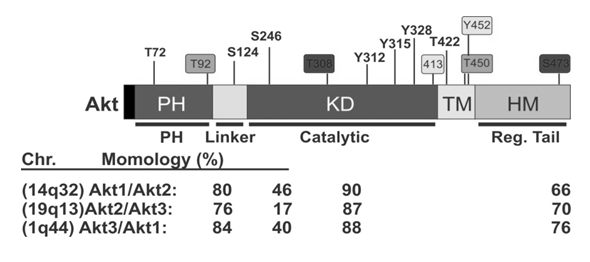
Figure 1a: Clustal format | Fasta format | MAFFT result.
Figure 1b shows the amino acid alignment of three isoforms (www.ncbi.org). It has been observed that there are major variations in amino acid sequences of the three isoforms that reside in their regulatory domain. Figure 2 shows the comparison of tertiary structure of three isoforms (http://zhanglab.ccmb.med.umich.edu/LOMETS). It may be seen, as expected from the major homologies between three isoforms that there is very little difference in their tertiary structure. Akt 3, however, shows an open loop in which Y452 is available for phosphorylation. The significance of this difference is discussed elsewhere in this report. It is now known that PIP 3 mediated phosphorylation of Akts by PDK 1 in kinase domains of Akt 1 - T308, Akt 2 - T306, and Akt 3 - T305 and by PDK 2 in the regulatory domains of Akt 1–S472, Akt 2–S478, and Akt 3–S474 [57]. It is also known that the phosphorylated isoforms are negatively regulated by PHiLPP1 and PHLiPP2. Phosphorylated threonine in the catalytic domain in each isoform is deregulated by SHiPs[58-60]. In figure 3 (a, b, c, d) are recorded results of amplification of Akt isoforms in the normal, Grade I, Grade II, and Grade III tissues of breast cancer. It may be noted that Akt 3 is conspicuously absent in the normal tissue while Akt 1 and Akt 2 show normal values though Akt 2 is higher than Akt 1. Progressively Akt 1 shows significant increase through Grade I, Grade II and Grade III. There is eye-catchingdifference in Akt 1 in grade I and Grade III. Akt 3 shows highly significant increase in Grade III compared to Grade I and Grade II. It shows elevation from the normal in Grade I and Grade III. The dynamics of three isoforms obtained in the study indicate that there is a considerable variation in three isoforms during the early and later stages in breast cancer tissue. A few samples of DISC and hyperplasia were also examined (data not shown in this report). From the data obtained on Akt isoforms the possible conceived role of three isoforms is presented in figure 4. In normal tissue only Akt 1 and Akt 2 are present. However, in the progression of breast cancer, there is a major event of transformation of epithelial cells to mesenchymal cells. For this at least in breast cancer we have observed that all three isoforms play a positive role in this transformation. As the tumor progresses, further invasive and metastasis stages, Akt 1 and Akt 3 seem to play dominant roles in metabolism, protein synthesis, proliferation, adhesion and growth[61,62].
Figure 1b: Structure and Akt isoform Domains. PH – Pleckstrin Homology Domain, KD – Kinase Domain, TM – Trans membrane region, HM – Hydrophobic Motif (Regulatory tail).
S – Serine, T – Threonine, Y – Tyrosine (Modified from Liao and Hung, 2010).
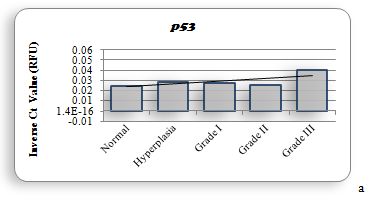
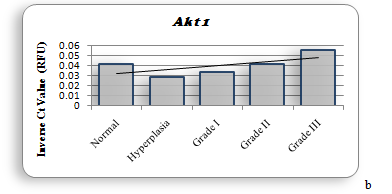
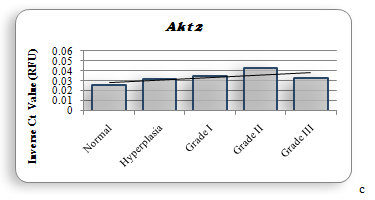
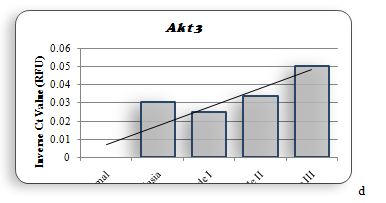
Figure 2
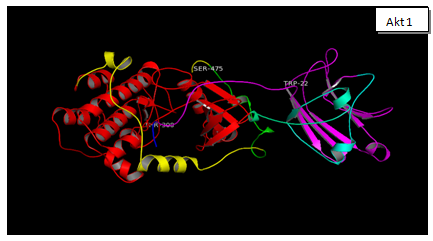
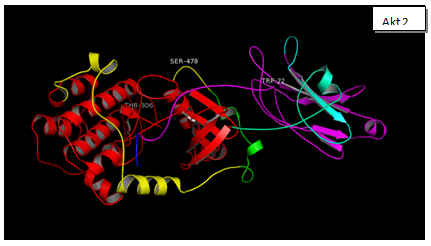
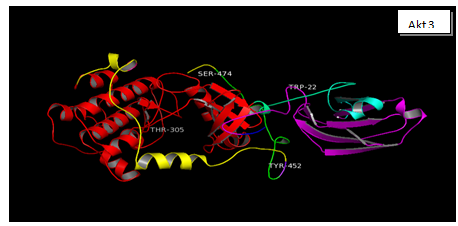
Figure 3: The three Akt Isoforms are presented the 3D structures of Akt 1, Akt 2 and Akt 3. After downloading the sequence of each isoform from NCBI site, gene accession number NM_005163.2 (Akt 1), NM_001626.3 (Akt 2), NM_181690.1 (Akt3). This was done to identify the possible phosphorylation sites in each isoform (www.phosida.org). It’s known that Akt 1, 2 and 3 are phosphorylated at threonine 308, 307 and 305; at serine 474,475 and 472. These Threonine sites are phosphorylated by PDK 1. However serine sites are phosphorylated by mToR C2, PDK 2 and ILK. These sites as may be seen in the 3D structure are exposed at the surface and are available for phosphorylation by the respective enzymes. In Akt 3 however additional sites: Ser413, Y 452 is also available for phosphorylation in the regulatory domain. The possible significance of these two sites has not been reported.
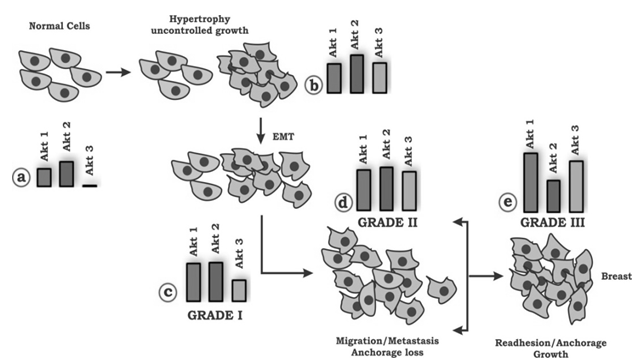
Figure 4: The figure demonstrates the possible conceived role played by three isoforms of Akt that is Akt 1, Akt 2 and Akt 3. It may be seen that in Normal tissue only Akt 1 and Akt 2 are present. However, during the progression of breast cancer, there is a major event of transformation of epithelial cells to mesenchymal cells. For this at least in breast cancer we have observed that all three isoforms play a positive role in this transformation. As the tumor progresses further into invasive and metastasis stages Akt 1 and Akt 3 seem to play dominant roles in metabolism, protein synthesis, proliferation, adhesion and growth.
Like the genes, 11 proteins associated with Akt-mToR pathway were estimated in normal and each grade of breast cancer tissues. The measurement of 11 phosphoproteins was simultaneously accomplished with Bioplex / Luminexx MAP based platform as described under materials and methods. The quantification was undertaken to find out whether these could serve as possible specific markers for various stages of breast cancer[63]. These proteins are mostly associated with metabolic requirements of proliferating cells. The estimated proteins included Akt (S473), mToR (S2448), GSK3α (S21), GSK3β (S9), p70 S6K (T412), RP S6 (S235/S236), IRS 1 (S312), PTEN (S380), TSC 2 (S939), IR (Y1162/Y1163), and IGF 1R (Y1135 / Y1136). The data obtained are recorded in table 1.
Table 1: Table represents values of various proteins ng/mL in normal and different Grades of breast cancer tissue.
| S.No | Names of Proteins | Normal | Hyperplasia | Grade I | Grade II | Grade III |
|---|---|---|---|---|---|---|
| 1 | pAkt (S473) | 19.08 | 25.81 | 24.50 | 23.00 | 19.75 |
| 2 | pmToR (S2448) | 19.08 | 20.66 | 20.08 | 18.25 | 18.50 |
| 3 | pGSK 3α (S21) | 14.58 | 16.47 | 15.42 | 18.42 | 16.25 |
| 4 | pGSK 3β (S9) | 9.42 | 12.00 | 12.25 | 10.75 | 7.25 |
| 5 | pp70S 6K (S412) | 37.83 | 21.09 | 41.33 | 38.00 | 40.25 |
| 6 | pRPS6 (S335/336) | 17.67 | 18.97 | 21.50 | 35.75 | 14.00 |
| 7 | pPTEN (S380) | 281.08 | 291.78 | 294.17 | 278.58 | 281.25 |
| 8 | pTSC2 (S939) | 15.00 | 23.84 | 19.69 | 16.50 | 24.50 |
| 9 | pIRS1 (S312) | 8.24 | 12.81 | 10.75 | 11.58 | 12.00 |
| 10 | pIR (Y1162/1163) | 24.67 | 29.41 | 23.50 | 26.08 | 24.50 |
| 11 | pIGF 1R (Y1135/1136) | 10.58 | 11.72 | 15.58 | 12.75 | 13.00 |
The dynamics of RCAS 1(Receptor associated Carcinoma Antigen on SiSo cells) like Akt isoforms was assayed in normal Grade I, Grade II and Grade III breast cancer tissues. RCAS 1 is strong indicator of immune evasion[31,34]. In all grades the relative amplification compared to normal tissue has been observed: more so in Grade II and Grade III. This indicates positive role of it in immune evasion, especially in the aggressive tissue. There is no significant difference in the amplification of any antiapoptotic genes (Bcl 2, BclxL and Mcl 1). This indicates that anti apoptotic role of these proteins remain unchanged. High levels of any of these Bcl 2 family proteins in Grade I, Grade II and Grade III tissues can serve as a major marker in prognosis of breast cancer after chemotherapy.
Discussion
In cancer, a large number of genes are known to be deregulated[64,66]. These include the genes of cellular proliferative RaS/MAPK pathway and survival Akt pathway[66]. Several lines of evidence suggest that hyperactivation of Akt signaling along with RaS transform normal cells to cancerous cells[67-69]. In view of these findings the present study was undertaken to obtain data on the dynamics of Akt isoforms and eleven proteins associated to Akt pathway in progression of breast cancer in various stages. In addition, three antiapoptotic genes namely Bcl 2, BclxL and Mcl 1 were quantified to assess their contribution and role in cellular proliferation. The three isoforms of Akt have shown close similarities in three dimensional structures as shown in figure 2. In spite of these similarities, specific functions have been assigned to each isoform especially in their modulations during the early and later stages of breast cancer[17,70]. Like others, we have also observed similar distribution of the three isoforms in normal, grade I, grade II and grade III breast cancer tissues. For instance, Akt 3 isoform is conspicuously absent in the normal tissue but is highly elevated in grade III tissues. This is in agreement with those reported by others[14,16,21,71-73]. Some recent reviews have discussed the functional specificity of Akt isoforms[2,74-75]. For instance, it has been suggested that Akt isoforms have relative importance in altered cells. It is substantiated that Akt 1 or Akt 2 or Akt 3 deficient mice, though viable, yet they have several deficiencies[76,77]. Another study Akt 2 knockout mice exhibited disorder in glucose metabolism (diabetes)[78,79]. This is accompanied by insulin resistance[80]. In Akt 3 deficient mice the brain size is reduced[16,81,82]. It has also been reported that Akt 2 levels are higher in 25% of breast carcinoma[24,83]. Compared to this, Akt 3 isoform is highly elevated in advanced stages of melanoma[71,72,84]. Furthermore, high level of Akt 3 activity is demonstrated in estrogen deficient breast cancer[35,85,86]. In essence, we have concluded from our data that differential constitutive elevation of Akt isoforms is responsible for initiation as well as progression of breast cancer as reported in other cancers. These results are in agreement with those reported by others[68,71,72,83].
A closer examination of our data reveals a relative progressive increase in Akt1, Akt 2 and Akt 3 in various grades of breast cancer tissue. This has been summarized in Normal, Grades I,II and III tissues (Figure 3). It has been documented that Akt 1 is a major isoform involved in proliferation of cancerous tissues[83,87,88]. Accordingly, the dominance of Akt 1 observed in this study is not surprising. This is further fortified by recent experimental evidence in which Akt 1 was down regulated by siRNA. This resulted in reduced proliferation[89,90]. In the same experiment, falls in the levels of Akt 1 resulted in reduced expression of Cyclin D1[85,91,92]. However, evidence to the contrary also exists. For instance, for purposes of proliferation, the role of Akt 2 cannot be minimized[79,87-95]. This was shown by enhancing the levels of Akt 2 in absence of Akt 1. In this case the proliferation was restored to the original rate of growth. It has been demonstrated that both Akt 1 and Akt 2 were required for in vitro and in vivo growth of human colon cancer cell line HCT 116 and DLD 1[96].
In view of our data which show high levels of Akt 1 and Akt 3 and reduced levels of Akt 2 in aggressive Grade III tissue, we are inclined to interpret that these changes are related to increase in mass and metastatic transformation which is characteristic of this stage[8]. In an excellent review by Sheng et al[15], it has been argued that metastasis must meet a minimum of nine requirements which include: relaxed proliferation, reduction in adhesion, a change in interaction with Extra Cellular Matrix (ECM), rapid turnover of altered fibroblasts, increase in the demand for nutrients and blood supply (angiogenesis), and focus of proliferation on metastatic sites followed by implantation[97,98]. It appears that in our case, based on quantitative gene analysis of 42 samples, the above conditions of metastasis are fulfilled primarily by Akt 1 and Akt 3 isoforms[99].
Though our data properly exhibits the dynamics of Akt isoforms in various grades of breast cancer tissue, yet it has to be explained whether in our case the increase of mRNA is due to; a) increased turnover of mRNA, b) genetic modifications, or c) gene amplification. It has been reported that genetic modifications are very rare in Akt[16]. Similarly, gene amplification of Akt has not been reported in breast cancer[20,100]. The differential elevation of mRNA of three isoforms leads to the conclusion that mRNA of each isoform is separately regulated. Furthermore, we have examined conformational differences in the three Akt isoforms using bioinformatics tools described under materials and methods. Interestingly enough, it has been reported that transplantation of PH domain of Akt 2 onto Akt 1 didn’t restore the function of proliferation characteristic of Akt 1[76,94]. We were particularly interested in identifying differences in 3D structure of PH domain for each isoform along with its phosphorylation sites[101,102]. 3D structures of the three isoforms, shown in figure 2, have been constructed using LOMETS (http://zhanglab.ccmb.med.umich.edu/LOMETS). It may be seen in Figure 2 that in addition to putative phosphorylation sites in the three isoforms, additional phosphorylation sites are available in Akt 3. The new sites of phosphorylation are S413 and Y452. Whether these sites are actually phosphorylated in the signaling process remains to be determined. It is interesting to note, as is evident from our data that in Grade III along with Akt 1 and Akt 3, kRaS is also elevated. This situation has been reported in literature in various types of cancer[35,103,104]. A model of Akt isoform dynamics has been reported earlier. The model is based on the modulation of only Akt 1 and Akt 2. In this report the model presents elevated Akt1 with depressed Akt 2. In EMT the situation is reversed and remains so in the metastatic stage[76,105]. In the data we are reporting in breast cancer patients in Pakistani females, we have observed the elevation of Akt 1 and Akt 3 in Grade III tissues. It is, however, important to point out that Akt 3 is not detected in the normal tissue (Figure 3). On these bases we can suggest that elevation in Akt 1 and Akt 3 in Grade III tissues can be taken as biomarkers for aggressive stages of breast cancer. This calls for use of specific inhibitors of these two Akt isoforms for the treatment of cancer[27,106].
In the Pakistani population we are reporting for the first time that Akt 3 isoform along with kRaS play a dominant role in the aggressive stage of breast cancer. It is also of interest that in this population Akt 2 is the first isoform to be elevated in hyperplasia. This is followed by elevation of Akt 1, Akt 2 and Akt 3 in subsequent stages (Figure 3 a, 3b, 3c).
In view of the well-known immunosuppressive role of RCAS 1[33,107-109] we were interested to study modulations of RCAS 1 because of its relationship with Akt 3 in various grades of breast cancer tissue. It has been documented that in tumor cells RCAS 1 expression plays an important role in evading the cancer cells from the immune system surveillance. This promotes progression, invasion and metastasis[27,34]. The antigen has been used as a prognostic marker in Normal, Hyperplasic and malignant uterine endometrium[3,109]. It has been identified as a possible factor in endocrine-immune interaction in breast cancer[33,110]. Elevated levels of this antigen have been reported in hepatocellular carcinoma[31], gastrointestinal tract, non-small cell lung carcinoma, gall bladder, breast, ovarian, endometrial and cervical cancers[33,111]. In our data presented in Figures 3f show the increase of RCAS 1 in Hyperplasia, Grade I, Grade II and Grade III tissues. This increase is directly related to high levels of Akt 3[84]. The antigen is also listed as EBAG 9 is known to bind estrogen receptor[112] thus activation of estrogen binding receptor. Gene suppression at transcription level and of immune system with MHC class 1 molecule on surface of tumor cells signifies various mechanisms that are in place in the cell[34,113]. In view of this information it appears that elevated levels of RCAS 1 in company with Akt 3 promote immune cell evasion, promotes survival, boost invasion and metastasis[108,114].
Similar conclusion could also be drawn with references to the levels of anti-apoptotic genes: Bcl 2, BclxL and Mcl 1 (Figures 3g, 3h, 3i). Our data also indicate that quantification of Bcl 2 family genes along with RCAS 1 could also be used as good prognostic markers as seen in prostate cancer[40,115,116]. Such a prognostic study must be accompanied by determining the levels of Akt 3 and anti apoptotic genes in the same tissue. This relationship of RCAS 1 with antiapoptotic genes and Akt 3 is being reported for the first time in Pakistani (Punjabi) population.
The proteins that were assayed for normal and different grades of breast cancer tissues are listed in table 1. Little information is available about the modulations of Akt and its substrate proteins in various grades of breast cancer. In our hands pAkt, pGSK 3β, pTSC 2, pIRS 1 and pIR can be conveniently used as biomarkers for Grade I and Grade II. However, we propose that in Grade III, pTSC 2, pIRS 1 and pIGF 1R quantification may serve as useful biomarkers. The proteins which are depressed in this grade include pGSK 3β and pRPS6. Comparable analytical data on these proteins in breast cancer tissue on various grades is not available in literature and are being reported for the first time. This could promote the process of grading breast cancer combined with other pathological information on Bloom-Richardson system[28]. It is of great interest to note that simultaneous analysis of gene reported in this study along with protein analysis could be of help in selecting future inhibitors for Akt and its substrates. The observation that levels of pAkt as well as pGSK 3α, pTSC 2, pIRS 1 and pIGF 1R are lowest in normal tissues (Table 1) may be of help in comparing of these proteins with their levels in various grades of cancerous tissues. pPTEN does not show any variation in the normal tissue as compared to its values in all other grades of cancer. pPTEN in active form dephosphorylates the PIP 3K stimulated phosphorylation of inositol at carbon 3[117,118]. The sustained levels of pPTEN in all tissue grades indicate its ineffectiveness to dephosphorylate PIP 3. We are therefore; suggest that elevated PTEN in the normal tissue is a mutated form of this gene (Data not shown). This has been previously reported in several studies[58,63,119-121].
The quantification of the proteins reported here, as expected, provided interesting information with regard to pIRS 1and pIGF 1R. Insulin is the ligand for the two receptors. These two proteins are elevated in all grades of breast cancer tissue[122]. This is indicative of increased glucose metabolism (energy) as is required during growth, proliferation and metastasis[23]. It has been previously reported that GSK 3α and GSK 3β were upregulated in normal prostate and in other cancers[123]. However, in breast cancer GSK 3β increases in Grade I and both GSK 3α and GSK 3β increase in Grade II. This is again indicative of the availability of energy through glucose metabolism[67,68,124].
We have estimated total mToR complex phosphorylated at S2448. mToR C2 phosphorylates Akt at S472 after its activation by pRheb[79,106,110]. Other substrates of mToR complex include STAT3, S6K1 (p70), 4E-BP1 and ULK1/ATG13[94,125]. In this way it promotes growth, proliferation, survival, angiogenesis and metabolic activities in various cancers[126,127]. Like others, we have observed increased levels of phospho p70 S6 kinase and pRPS6. Alternatively, RPS6 is also phosphorylated by p90 (RSK). In that case it inhibits the selective transcription of various other genes[94,128] making significant changes in normal cell signaling cascades. This is more evident from phosphorylated RPS6 that promotes growth factors and tumor promoting agents[128].
Conclusion
In essence, our genomic and proteomic data provide substantial evidence that Akt 1 and Akt 3 provide signals for proliferation of tumor (size) and invasion – angiogenesis. Furthermore, the elevated levels of pIR, pIGF 1R, pIRS 1 in glucose metabolism accompanied by phosphorylation of GSK 3α and GSK 3β provide higher energy source for growth, survival, proliferation and angiogenesis. The subtle dynamics of genes and proteins reported in this study may be of some help in selection of therapeutic molecules for breast cancer.
Acknowledgement: I thank University of Lahore, Lahore 54000 for providing financial support for Milliplex Akt-mToR kit. I also acknowledge expert statistical work by MsRabailAlam, IMBB, University of Lahore for statistical layout / analysis and MsSumairaShaheen, CRiMM, University of Lahore for critical manuscript evaluation / formatting.
References
1. Hanif, M., Zaidi, P., Kamal, S., et al. Institutional based Cancer incidence in a local population in Pakistan: nine year data analysis. (2009) Asian Pac J Cancer Prev 10(2): 227-230.
Pubmed||Crossref||Others
2. Bhurgri, Y., Bhurgri, A., Nishter, S., et al. Pakistan country profile of cancer and cancer control 1995-2004. (2006) J Pak Med Assoc 56(3): 124-130.
Pubmed||Crossref||Others
3. Mamoon, N., Sharif, M.A., Mushtaq, S., et al. Breast carcinoma over three decades in northern Pakistan|| Are we getting anywhere|| (2009) JPMA 59(12): 835-838.
Pubmed||Crossref||Others
4. DeSantis, C., Siegel, R., Bandi, P., et al. Breast Cancer Statistics, 2011. (2011) CA Cancer J Clin 61(6): 409-418.
Pubmed||Crossref||Others
5. Women Health Problems. Breast Cancer. 2009.
Pubmed||Crossref||Others
6. Fatima, N., Zaman, M.U., Maqbool, A., et al. Lower incidence but more aggressive behavior of right sided breast cancer in Pakistani women: does right deserve more respect||. (2013) Asian Pac J Cancer Prev 14(1): 43-45.
Pubmed||Crossref||Others
7. Parveen, S., Sarwar, G., Khuwaja, M. Carcinoma of breast, pattern and presentation in developing countries. (2011) Pak J Surg 27(4): 246-249.
Pubmed||Crossref||Others
8. Cooper, C.L., Karim, R.Z., Selinger, C., et al. Molecular alterations in metaplastic breast carcinoma. (2013) J ClinPathol 66(6): 522-528.
Pubmed||Crossref||Others
9. Mamoon, N., Hussan, U., Mushtaq, S. Breast carcinoma in young women age 30 or less in northern Pakistan – the armed forces institute of Pathology experience. (2009) Asian Pac J Cancer Prev 10(6): 1079-1082.
Pubmed||Crossref||Others
10. Carnero, A. The PKB/Akt pathway in cancer. (2010) Cur Pharmaceut Design 16(1): 34-44.
Pubmed||Crossref||Others
11. Lemmon, M.A., Schlessinger, J. Cell signaling by receptor tyrosine kinases. (2010) Cell 141(7): 1117-1134.
Pubmed||Crossref||Others
12. Dillon, R.L., Muller, W.J. Distinct biological roles for the Akt family in mammary tumor progression. (2010) Cancer Res 70(11): 4260-4264.
Pubmed||Crossref||Others
13. Diez, H., Garrido, J.J., Wandosell, F. Specific roles of Akt isoforms in apoptosis and axon growth regulation in neurons. (2012) PLoS One 7(4): e32715.
Pubmed||Crossref||Others
14. Bellacosa, A., Testa, J.R., Larue, L. A portrait of Akt kinases: human cancer and animal models depict a family with strong individualities. (2004) Cancer Biol Ther 3(3): 268-275.
Pubmed||Crossref||Others
15. Sheng, S., Qiao, M., Pardee, A.B. Metastasis and AKT activation. (2009) J Cell Physiol 218(3): 451-454.
Pubmed||Crossref||Others
16. Bellacosa, A., Larue, L. PI3K/Akt Pathway and the Epithelial-Mesenchymal Transition. (2010) Springer Sci, USA.
Pubmed||Crossref||Others
17. Hemmings, B.A., Restuccia, D.F. PI3K-PKB / Akt pathway. (2012) Cold Spring Harbor Perspect Biol 4(9): a011189.
Pubmed||Crossref||Others
18. Hutchinson. J., Jin, J., Cardiff, R.D., et al. Activation of Akt (protein kinase B), in mammary epithelium provides a critical cell survival signal required for tumor progression. (2001) Mol Cellular Biol 21(6): 2203-2212.
Pubmed||Crossref||Others
19. Peng, Z., Weber, J.C., Han, Z. Dichotomy effects of Akt signalling in breast cancer. (2012) Mol Cancer 11: 61.
Pubmed||Crossref||Others
20. Brugge, J.S., Hung, M-C., Mills, G.B. A New Mutational Aktivation in the PI3K Pathway. (2007) Cancer Cell 12(2): 104-107.
Pubmed||Crossref||Others
21. Calvo, M.B., Fernandez, V.B., Villaamil, V.M., et al. Biology of BMP signalling and cancer. (2009) Clin Transl Oncol 11(3): 126-137.
Pubmed||Crossref||Others
22. Davies, M.A. Regulation, role, and targeting of Akt in cancer. (2011) J Clin Oncol 29(35): 4715-4717.
Pubmed||Crossref||Others
23. Cairns, R.A., Harris, I.S., Mak, T.W. Regulation of cancer cell metabolism. (2011) Nat Rev Cancer 11(2): 85-95.
Pubmed||Crossref||Others
24. Sung, J.S., Park, K.H., Kim, S.T., et al. Discovery and evaluation of polymorphisms in the Akt 2 and Akt 3 promoter regions for risk of Korean lung cancer. (2012) Genomics Inform 10(3): 167-174.
Pubmed||Crossref||Others
25. Do, H., Salemi, R., Murone, C., et al. Rarity of Akt 1 and Akt 3 E17K mutations in squamous cell carcinoma of lung. (2010) Cell Cycle 9(21): 4411-4412.
Pubmed||Crossref||Others
26. Brunelle, J.K., Letai, A. Control of mitochondrial apoptosis by the Bcl 2 family. (2009) J Cell Sci 122(4): 437-441.
Pubmed||Crossref||Others
27. Chin, Y.R., Toker, A. Akt isoform specific signaling in breast cancer: Uncovering an anti-migratory role for paladin. (2011) Cell Adh Migr 5(3): 211-214.
Pubmed||Crossref||Others
28. Bloom, H.J.G., Richardson, W.W. Histological grading and prognosis in breast cancer: a study of 1409 cases of which 359 have been followed for 15 years. (1957) Br J Cancer 11(3): 359.
Pubmed||Crossref||Others
29. Carpten, J.D., Faber, A.L., Horn, C., et al. A transforming mutation in the pleckstrin homology domain of Akt 1 in cancer. (2007) Nature 448(7152): 439-444.
Pubmed||Crossref||Others
30. Migita, T., Inoue, S. Implications of the Golgi apparatus in prostate cancer. (2012) Intl J Biochem Cell Biol 44(11): 1872-1876.
Pubmed||Crossref||Others
31. Jozwicki, W., Windorbska, W., Brozyna, A.A., et al. The analysis of receptor-binding cancer antigen expressed on SiSo cells (RCAS 1); immunoreactivity within the microenvironment of the ovarian cancer lesion relative to the applied therapeutic strategy. (2011) Cell Tissue Res 345(3): 405-414.
Pubmed||Crossref||Others
32. Grabinski, N., Bartkowiak, K., Grupp, K., et al. Distinct functional roles of Akt isoforms for proliferation, survival, migration and EGF-mediated signalling in lung cancer derived disseminated tumor cells. (2011) Cell Signal 23(12): 1952-1960.
Pubmed||Crossref||Others
33. Sonoda, K. RCAS 1 is a promising therapeutic target against cancer: its multifunctional bioactivities and clinical significance. (2012) Expert Rev Obst Gynaecol 7(3): 261-267.
Pubmed||Crossref||Others
34. Nishinakagawa, T., Fujii. S., Nozaki, T., et al. Analysis of cell cycle arrest and apoptosis induced by RCAS 1 expression. (2010) Int J Mol Med 25(5):717-722.
Pubmed||Crossref||Others
35. Hanrahan, A.J., Schultz, N., Westfal, M.L., et al. Genomic complexity and Akt dependence in serous ovarian cancer. (2012) Cancer Discov 2(1): 56-67.
Pubmed||Crossref||Others
36. Chipuk, J.E., Moldoveanu, T., Llambi, F., et al. The Bcl 2 family reunion. (2010) Mol Cell 37(3): 299-310.
Pubmed||Crossref||Others
37. Renault, T.T., Chipuk, J.E. Getting away with murder: how does the Bcl 2 family of proteins kill with immunity|| (2013) Ann NY Acad Sci 1285: 59-79.
Pubmed||Crossref||Others
38. Bolesta, E., Pfannenstiel, L.W., Demelash, A., et al. Inhibition of Mcl 1 promotes senescence in cancer cells: implications for preventing tumor growth and chemotherapy resistance. (2012) Mol Cell Biol 32(10): 1879-1892.
Pubmed||Crossref||Others
39. Kazi, A., Sun, J., Doi, K., et al. The BH3 alpha-helical mimic BH3-M6 disrupts BclxL, Bcl-2, and Mcl 1 protein-protein interactions with Bax, Bak, Bad, or Bim and induces apoptosis in a Bax- and Bim-dependent manner. (2011) J Biol Chem 286(11): 9382-9392.
Pubmed||Crossref||Others
40. Dewson, G., Kluck, R.M. Bcl 2 family regulated apoptosis in health and disease. (2010) Cell Health Cytoskelton 2010: 9-22.
Pubmed||Crossref||Others
41. Flusberg, D.A., Numaguchi, Y., Ingber, D.E. Cooperative Control of Akt phosphorylation, Bcl 2 expression, and apoptosis by cytoskeletal microfilaments and microtubules in capillary endothelial Cells. (2001) Mol Biol Cell 12(10): 3087-3094.
Pubmed||Crossref||Others
42. Okaro, A.C., Deery, A.R., Hutchins, R.R. et al. The expression of antiapoptotic proteins Bcl 2, BclxL, and Mcl 1 in benign, dysplastic, and malignant biliary epithelium. (2001) J Clin Pathol 54(12): 927-932.
Pubmed||Crossref||Others
43. Dalafave, D., Presto, G. Inhibition of antiapoptotic BclxL, Bcl 2, and Mcl 1 proteins by small molecule mimetics. (2010) Cancer Inform 9: 169-177.
Pubmed||Crossref||Others
44. Kallel-Bayoudh, I., Hassen, H.B., Khabir, A., et al. Bcl 2 expression and triple negative profile in breast carcinoma. (2011) Medical Oncol 28(Suppl 1): S55-S61.
Pubmed||Crossref||Others
45. Shamas-Din, A., Kale, J., Leber, B., et al. Mechanisms of action of Bcl 2 family proteins. (2013) Cold Spring Harbor Perspect Biol 5(4): a008714.
Pubmed||Crossref||Others
46. Huang, J., Manning, B.D. A complex interplay between Akt, TSC2, and the two mTOR complexes. (2009) Biochem Soc Trans 37(1): 217-222.
Pubmed||Crossref||Others
47. Colo, J.L., Macintyre, A.N., et al. Akt dependent glucose metabolism promotes Mcl 1 synthesis to maintain cell survival and resistance to Bcl 2 inhibition. (2011) Cancer Res 71(15): 5204-5213.
Pubmed||Crossref||Others
48. Gupta, M., Hendrickson, A.E., et al. Dual mTORC1 / mTORC2 inhibition diminishes Akt activation and induces Puma dependent apoptosis in lymphoid malignancies. (2012) Blood 119(2): 476-487.
Pubmed||Crossref||Others
49. O’Driscoll, L., Cronin, D., et al. Expression and prognostic relevance of Mcl 1 in breast cancer. (2004) Anticancer Res 24(2A): 473-482.
Pubmed||Crossref||Others
50. Altomare, D.A., Testa, J.R. Perturbations of the Akt signaling pathway in human cancer. (2005) Oncogene 24(50): 7455-7464.
Pubmed||Crossref||Others
51. Shao, Y., Aplin, A.E. Akt 3 - mediated resistance to apoptosis in BRaf-targeted melanoma cells. (2010) Cancer Res 70(16): 6670-6681.
Pubmed||Crossref||Others
52. Kirkegaard, T., Edwards, J., et al. Molecular alterations in Akt 1, Akt 2 and Akt 3 detected in breast and prostatic cancer by FISH. (2010) Histopathol 56(2): 203-211.
Pubmed||Crossref||Others
53. Farrell, R.E. RNA Methodologies. (2010) Academic Press USA.
Pubmed||Crossref||Others
54. Farragher, S.M., Tanney, A., et al. RNA expression analysis from formalin fixed paraffin embedded tissues. (2008) Histochem Cell Bio 130(3): 435-445.
Pubmed||Crossref||Others
55. Dey-Guha, I., Wolfer, A., Yeh, A.C., et al. Asymmetric cancer cell division regulated by Akt. (2011) Proc Natl Acad Sci 108(31): 12845-12850.
Pubmed||Crossref||Others
56. Chua, B.T., Gallego-Ortega, D., Ramirez-de-Molina, A., et al. Regulation of Akt (S473) phosphorylation by choline kinase in breast carcinoma cells. (2009) Mol Cancer 8: 131.
Pubmed||Crossref||Others
57. Novak, B., Kapuy, O., Domingo-Sananes, M.R., et al. Regulated protein kinases and phosphatases in cell cycle decisions. (2010) Cur Opin Cell Biol 22(6): 801-808.
Pubmed||Crossref||Others
58. Chan, T.O., Zhang, J., Rodeck, U., et al. Resistance of Akt kinases to dephosphorylation through ATP dependent conformational plasticity. (2011) Proc Natl Acad 108(46): 1120-1127.
Pubmed||Crossref||Others
59. Sun, H., Wang, Y. Novel Serine / Threonine protein phosphatases in cell death regulation. (2012) Physiol 27(1): 43-52.
Pubmed||Crossref||Others
60. Haslinger, P., Haider, S., Sonderegger, S., et al. Akt isoforms 1 and 3 regulate basal and epidermal growth factor-stimulated SGHPL-5 trophoblast cell migration in humans. (2013) Biol Reprod 88(3): 54.
Pubmed||Crossref||Others
61. Liang, Y., Liu, J., Feng, Z. The regulation of cellular metabolism by tumor suppressor p53. (2013) Cell Biosci 3(1): 9.
PubMed||Crossref||Others
62. McCrea, H.J., De Camilli, P. Mutations in Phosphoinositide Metabolizing Enzymes and Human Disease. (2009) Physiol 24(1): 8-16.
PubMed||Crossref||Others
63. Agathocleous, M., Harris, W.A. Metabolism in physiological cell proliferation and differentiation. (2013) Trends Cell Biol 23(10): 484-492.
PubMed||Crossref||Others
64. Brognard, J., Hunter, T. Protein kinase signaling networks in cancer. (2011) Curr Opin Genet Dev 21(1): 4-11.
PubMed||Crossref||Others
65. Castellano, E., Downward, J. RAS interaction with PI3K: more than just another effector pathway. (2011) Genes Cancer 2(3): 261-274.
PubMed||Crossref||Others
66. Cecconi, S., Mauro, A., Cellini, V., et al. The role of Akt signalling in the mammalian ovary. (2012) Intl J Develop Biol 56(10-12): 809-817.
PubMed||Crossref||Others
67. Cristiano, B.E., Chan, J.C., Hannan, K.M., et al. A specific role for Akt 3 in the genesis of ovarian cancer through modulation of G2 – M phase transition. (2006) Cancer Res 66(24): 11718-11725.
PubMed||Crossref||Others
68. Denayer, E., de Ravel, T., Legius, E. Clinical and molecular aspects of RAS related disorders. (2008) J Med Genet 45(11): 695-703.
PubMed||Crossref||Others
69. Virtakoivu, R., Pellinen, T., Rantala, J.K., et al. Distinct roles of Akt isoforms in regulating β1-integrin activity, migration, and invasion in prostate cancer. (2012) Mol Biol Cell 23(17): 3357-3369.
PubMed||Crossref||Others
70. Xu, N., Lao, Y., Zhang, Y., et al. Akt: a double-edged sword in cell proliferation and genome stability. (2012) J Oncol 2012: 15.
PubMed||Crossref||Others
71. Wickendon, J.A., Watson, C.J. Key signalling nodes in mammary gland development and cancer, signalling downstream of PI3 kinase in mammary epithelium: a play in 3 Akts. (2010) Cancer Res 12(2): 202.
PubMed||Crossref||Others
72. Gonzalez, E., McGraw, T.E. The Akt kinases: isoform specificity in metabolism and cancer. (2009) Cell Cycle 8(16): 2502-2508.
PubMed||Crossref||Others
73. Fortier, A.M., Asselin, E., Cadrin, M. Functional specificity of Akt isoforms in cancer progression. (2011) Bio Mol Concepts 2(1-2): 1-11.
PubMed||Crossref||Others
74. Toker, A. Achieving specificity in Akt signaling in cancer. (2012) Adv Biol Regul 52(1): 78-87.
PubMed||Crossref||Others
75. Fortier, A.M., Van, Themsche, C., Asselin, E., et al. Akt isoforms regulate intermediate filament protein levels in epithelial carcinoma cells. (2010) FEBS Lett 584(5): 984-988.
PubMed||Crossref||Others
76. Yang, W-L., Wu, C-Y., Wu, J., et al. Regulation of Akt signaling activation by ubiquitination. (2010) Cell Cycle 9(3): 487-497.
PubMed||Crossref||Others
77. Buzzi, F., Xu, L., Zuellig, R.A., et al. Differential effects of protein kinase B/Akt isoforms on glucose homeostasis and islet mass. (2010) Mol Cell Biol 30(3): 601-612.
PubMed||Crossref||Others
78. Chau, N.M., Ashcroft, M. Akt 2: a role in breast cancer metastasis. (2004) Br Cancer Res 6(1): 55-57.
PubMed||Crossref||Others
79. Gonzalez, E., McGraw, T.E. Insulin modulated Akt subcellular localization determines Akt isoform specific signaling. (2009) Proc Natl Acad Sci U S A 106(17): 7004-7009.
PubMed||Crossref||Others
80. Chen, Y.Y., Li, C.F., Yeh, C.H., et al. Interleukin-19 in breast cancer. (2013) Clinical Develop Immunol 2013: 9.
PubMed||Crossref||Others
81. Toker, A. Achieving specificity in Akt signaling in cancer. (2012) Adv Biol Regul 52(1): 78-87.
PubMed||Crossref||Others
82. Duan, W. mRNA expression of three isoforms of Akt in different human cancer cell lines. (2011) Biotech. PubMed||Crossref||Others
83. Liby, T.A., Spyropoulos, P., Buff, L.H., et al. Akt 3 controls vascular endothelial growth factor secretion and angiogenesis in ovarian cancer cells. (2012) Int J Cancer 130(3): 532-543.
PubMed||Crossref||Others
84. Liao, Y., Hung, M-C. Physiological regulation of Akt activity and stability. (2010) Am J Transl Res 2(1): 19-42.
PubMed||Crossref||Others
85. Nakatani, K., Thompson, D.A., Barthel, A. Upregulation of Akt 3 in estrogen receptor deficient breast cancers and androgen-independent prostate cancer lines. (1999) J Biol Chem 274(31): 21528-21532.
PubMed||Crossref||Others
86. Lee, R.S., House, C.M., Cristiano, B.E., et al. Relative expression levels rather than specific activity plays the major role in determining in vivo Akt isoform substrate specificity. (2011) Enzyme Res 2011: 720985.
PubMed||Crossref||Others
87. Devaney, J.M., Gordish-Dressman, H., Harmon, B.T., et al. Akt 1 polymorphisms are associated with risk for metabolic syndrome. (2011) Hum Genet 129(2): 129-139.
PubMed||Crossref||Others
88. Small, E.M., O’Rourke, J.R., Moresi, V., et al. Regulation of PI3-kinase / Akt signaling by muscle-enriched microRNA-486. (2010) Proc Natl Acad Sci U S A 107(9): 4218-4223.
PubMed||Crossref||Others
89. Meng, Z., Lou, S., Tan, J. et al. Nuclear factor-kappa B inhibition can enhance therapeutic efficacy of 131I on the in vivo management of differentiated thyroid cancer. (2012) Life sci 91(23-24): 1236-1241.
PubMed||Crossref||Others
90. Chan, T.O., Pascal, J.M., Armen, R.S. et al. Autoregulation of kinase dephosphorylation by ATP binding in AGC protein kinases. (2012) Cell Cycle 11(3): 475-478.
PubMed||Crossref||Others
91. Dey-Guha, I., Wolfer, A., Yeh, A.C., et al. Asymmetric cancer cell division regulated by Akt. (2011) PNAS 108(31): 12845-12850.
PubMed||Crossref||Others
92. Santi, S.A., Lee, H. Ablation of Akt 2 induces autophagy through cell cycle arrest, the down regulation of p70S6K, and the deregulation of mitochondria in MDA-MB231 cells. (2011) PLoS One 6(1): e14614.
PubMed||Crossref||Others
93. Liu, Y., Yang, G., Bu, X., et al. Cell-type-specific regulation of raft-associated Akt signaling. (2011) Cell Death Dis 2: e145.
PubMed||Crossref||Others
94. Sheng, S., Qiao, M., Pardee, A.B. Metastasis and AKT activation. (2009) J cell physiol 218(3): 451-454.
PubMed||Crossref||Others
95. Los, M., Maddika, S., Erb, B., et al. Switching Akt: from survival signaling to deadly response. (2009) BioEssays 31(5): 492-495.
PubMed|| Crossref ||Others
96. Bluff, J.E., Brown, N.J., Reed, M.W., et al. Tissue factor, angiogenesis and tumor progression. (2008) Br Cancer Res 10(2): 204.
PubMed||Crossref||Others
97. Schultze, S.M., Jensen, J., Hemmings, B.A., et al. Promiscuous affairs of PKB / AKT isoforms in metabolism. (2011) Arch Physiol Biochem 117(2): 70-77.
PubMed||Crossref||Others
98. Shiojima, I. Role of Akt Signaling in Vascular Homeostasis and Angiogenesis. (2002) Circ Res 90(12): 1243-1250.
PubMed||Crossref||Others
99. Ellsworth, R.E., Decewicz, D.J., Shriver, C.D., et al. Breast cancer in the personal genomics era. (2010) Cur Genomics 11(3): 146-161.
PubMed||Crossref||Others
100. Gao, X., Lowry, P.R., Zhou, X., et al. PI3K / Akt signaling requires spatial compartmentalization in plasma membrane microdomains. (2011) Proc Natl Acad Sci U S A 108(35): 14509-14514.
PubMed||Crossref||Others
101. Wullschleger, S., Sakamoto, K., Johnstone, L., et al. How moderate changes in Akt T-loop phosphorylation impact on tumorigenesis and insulin resistance. (2011) Dis Model Mech 4(1): 95-103.
PubMed||Crossref||Others
102. Hanker, A.B., Der, C.J. The roles of RaS family small GTPases in Breast Cancer. (2010) Cell Signal Coll 2763-2772.
PubMed||Crossref||Others
103. Omerovic, J., Laude, A.J., Prior, I.A. Ras proteins: paradigms for compartmentalized and isoform-specific signalling. (2007) Cellular Mol Life Sci 64(19-20): 2575-2589.
PubMed||Crossref||Others
104. Polyak, K., Weinberg, R.A. Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. (2009) Nat Rev Cancer 9(4): 265-273.
PubMed||Crossref||Others
105. Memmott, R.M., Dennis, P.A. Akt-dependent and -independent mechanisms of mTOR regulation in cancer. (2009) Cellular Signalling 21(5): 656-664.
PubMed||Crossref||Others
106. Zhang, Y., Liu, X., Zhang, J., et al. The expression and clinical significance of PI3K, pAkt and VEGF in colon cancer. (2012) Oncol Lett 4(4): 763-766.
PubMed||Crossref||Others
107. Dutsch-Wicherek, M., Lazar, A., Tomaszewska, R. The involvement of RCAS 1 in creating a suppressive tumor microenvironment in patients with salivary gland Adenocarcinoma. (2010) Cancer Microenviron 4(1): 13-21.
PubMed||Crossref||Others
108. Adamek, D., Radwa||ska, E., Gajda, M. Expression of RCAS 1 protein in microglia/macrophages accompanying brain tumors, an immunofluorescence study. (2009) Folia Neuropathol 47(3): 240-246.
PubMed||Crossref||Others
109. Du, K., Tsichlis, P.N. Regulation of the Akt kinase by interacting proteins. (2005) Oncogene 24(50): 7401-7409.
PubMed||Crossref||Others
110. Srivastava, K., Srivastava, A., Mittal, B. Potential biomarkers in gallbladder cancer: present status and future directions. (2013) Biomarkers 18(1): 1-9.
PubMed||Crossref||Others
111. Kino, T., Clhrousos, G.P. Tumor associated, Estrogen Receptor related Antigen EBAG9: linking intracellular vesicle trafficking, immune homeostasis and malignancy. (2009) Mol Interv 9(6): 294-298.
PubMed||Crossref||Others
112. Maddocks, O.D., Vousden, K.H. Metabolic regulation by p53. (2011) J Mol Med 89(3): 237-245.
PubMed||Crossref||Others
113. Dutsch-Wicherek, M., Tomaszewska, R., Lazar, A., et al. The associations between RCAS1 expression in laryngeal and pharyngeal cancer and its healthy stroma with cancer relapse. (2009) BMC Cancer 9: 35.
PubMed||Crossref||Others
114. Brunelle, J.K., Letai, A. Control of mitochondrial apoptosis by the Bcl 2 family. (2009) J Cell Sci 122(Pt 4): 437-441.
PubMed||Crossref||Others
115. Kumagai, J., Urano, T., Ogushi, T., et al. EBAG 9 is a tumor||promoting and prognostic factor for bladder cancer. (2009) Intl J Cancer 124(4): 799-805.
PubMed||Crossref||Others
116. Adams, J.R., Schachter, N.F., Liu, J.C., et al. Elevated PI3K signaling drives multiple breast cancer subtypes. (2011) Oncotarget 2(6): 435-447.
PubMed||Crossref||Others
117. Datta, S.R., Dudek, H., Tao, X., et al. Akt phosphorylation of BAD couples survival signals to the cell intrinsic death machinery. (1997) Cell 91(2): 231-241.
PubMed||Crossref||Others
118. Longuespee, R., Boyon, C., Desmons, A., et al. Ovarian cancer molecular pathology. (2012) Cancer Metastasis Rev 31(3-4): 713-732.
PubMed||Crossref||Others
119. Georgescu, M.-M. PTEN Tumor Suppressor Network in PI3K-Akt Pathway Control. (2010) Genes Cancer 1(12): 1170-1177.
PubMed||Crossref||Others
120. Jones, R.G., Thompson, C.B. Tumor suppressors and cell metabolism: a recipe for cancer growth. (2009) Genes Dev 23(5): 537-548.
PubMed||Crossref||Others
121. Djiogue, S., Kamdje, A.H.N., Vecchio, L. Insulin resistance and cancer: the role of insulin and IGFs. (2013) Endocr Relat Cancer 20(1): R1-R17.
PubMed||Crossref||Others
122. Darrington, R.S., Campa, V.M., Walker, M.M., et al. Distinct expression and activity of GSK 3α and GSK 3β in prostate cancer. (2012) Int J Cancer 131(6): E872-883.
PubMed||Crossref||Others
123. Matheny, R.W. Jr., Adamo, M.L. Current perspectives on Akt Akt-ivation and Akt-ions. (2009) Exp Biol Med 234(11): 1264-1270.
PubMed||Crossref||Others
124. Azab, S.S. Targeting the mToR signalling pathways in breast cancer: more than the rapalogs. (2013) J Biochem Pharmacological Res 1(2): 75-83.
PubMed||Crossref||Others
125. Cidado, J., Park, B.H. Targeting the PI3K / Akt / mTOR pathway for breast cancer therapy. (2012) J Mammary Gland Biol Neoplasia 17(3-4): 205-216.
PubMed||Crossref||Others
126. Moritz, A., Li, Y., Guo, A. Akt – RSK - S6 kinase signaling networks activated by oncogenic receptor tyrosine kinases. (2010) Sci Signal 3(136): ra64.
PubMed||Crossref||Others
127. Blanco-Aparicio, C., Canamero, M., Cecilia, Y., et al. Exploring the gain of function contribution of Akt to mammary tumorigenesis in mouse models. (2010) PLoS One 5(2): e9305.
PubMed||Crossref||Others
128. Dummler, B., Tschopp, O., Hynx, D. Life with a single isoform of Akt: mice lacking Akt 2 and Akt 3 are viable but display impaired glucose homeostasis and growth deficiencies. (2006) Mol Cell Biol 26(21): 8042-8051.
PubMed||Crossref||Others










